
Melanoma is a type of skin cancer. It arises from specialized pigmented cells in our body known as melanocytes that are responsible for the production of melanin (a pigment responsible for skin and hair color). Because most melanoma cells still make melanin, melanoma tumors are usually brown or black. It accounts for 4% of all skin cancers; however, it is responsible for the largest numbers of skin cancer related death in the world. In the US, according to the national cancer institute, estimated new cases and deaths from melanoma in 2013 would be 76,690 and 9,480 respectively.
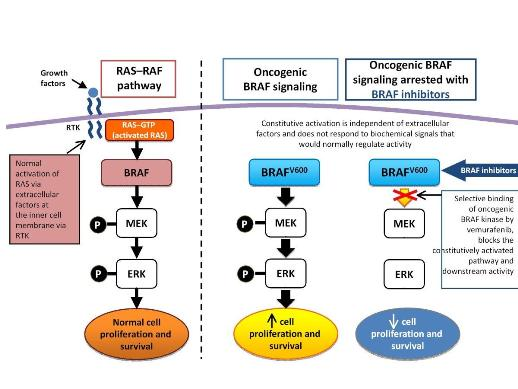
Several studies using molecular profiling and genomic sequencing have shown that melanoma is a disease of a heterogeneous group of tumors, and its progression is driven by specific oncogenic mutations. In 2002, Davies et al. first reported the presence of B-RAF somatic missense mutations in 66% of malignant melanomas. RAF (Rapidly growing Fibrosarcoma) protein is a serine/thereonine kinase. Three members of this kinase family are A-RAF, B-RAF, and C-RAF. These serine/threonine protein kinases, downstream of the membrane-bound small G protein RAS, are components of the mitogen activtated protein kinase (MAPK) signal transduction pathway. With closely overlapping functions, all members of the RAF family are associated with the activation of the MAPK pathway. Activation of the MAPK pathway has been associated with uncontrolled growth and drug resistance in several tumors. Researchers have identified over 50 distinct mutations in the B-RAF gene so far. However, most of these mutations are extremely rare. The most common mutation in melanoma, accounting for 90% of all B-RAF mutations, is the V600E mutation that occurs as a result of substitution of amino acid valine (V) to glutamic acid (E) at codon 600. Approximately 50% of melanomas harbor the V600E B-RAF mutation, while other mutations observed in melanomas are usually associated with the activation of N-RAS and c-KIT.
growing Fibrosarcoma) protein is a serine/thereonine kinase. Three members of this kinase family are A-RAF, B-RAF, and C-RAF. These serine/threonine protein kinases, downstream of the membrane-bound small G protein RAS, are components of the mitogen activtated protein kinase (MAPK) signal transduction pathway. With closely overlapping functions, all members of the RAF family are associated with the activation of the MAPK pathway. Activation of the MAPK pathway has been associated with uncontrolled growth and drug resistance in several tumors. Researchers have identified over 50 distinct mutations in the B-RAF gene so far. However, most of these mutations are extremely rare. The most common mutation in melanoma, accounting for 90% of all B-RAF mutations, is the V600E mutation that occurs as a result of substitution of amino acid valine (V) to glutamic acid (E) at codon 600. Approximately 50% of melanomas harbor the V600E B-RAF mutation, while other mutations observed in melanomas are usually associated with the activation of N-RAS and c-KIT.
Several studies reported association of the V600E B-RAF mutation with the progression of melanoma. In a pre-clinical study Smalley et al. (2010) observed tumor formation in immunocompromised mice following introduction of mutant B-RAF in melanocytes. Inversely, in their study, Smalley et al. also observed that inhibition of mutated B-RAF using RNA-interference resulted in tumor cell death. In addition, several other studies reported that inhibition of V600E mutant B-RAF prevents melanoma cell proliferation, induces apoptosis (programmed cell death), and also blocks melanoma xenograft growth in vivo. Even though many studies suggested that V600E B-RAF mutation may not be sufficient alone for melanoma induction, a wealth of evidence demonstrated that mutated B-RAF is necessary for the maintenance and progression of melanoma in human. Therefore, mutated B-RAF represents a therapeutic target in melanoma, which is why several B-RAF kinase inhibitors have already been developed. Sorafenib was the first B-RAF inhibitor studied in melanoma patients. In addition, vemurafenib (Zelboraf) and dabrafenib (GSK2118436) were also studied in melanoma patients with V600E B-RAF mutations. In 2011 vemurafenib received FDA approval for the treatment of melanoma patients harboring the V600E B-RAF mutation. In clinical trials, in which patients were undergoing treatment with vemurafenib, the drug reduced risk of death by 63% and risk of progression by 74%.
At present several clinical trials also evaluate clinical efficacy of vemurafenib in combination with leflunomide (antirheumatic drug), GDC-0973 (MEK inhibitor), and metformin (antidiabetic drug). In addition, several other drugs targeting B-RAF and its downstream pathway are also in development. Therefore, further improvements can be expected in this personalized and targeted therapy in melanoma.
References:
1. Ascierto, P. A., Kirkwood, J. M., Grob, J. J., Simeone, E., Grimaldi, A. M., Maio, M., Palmieri, G., Testori, A., Marincola, F. M., and Mozzillo, N. (2012). The role of BRAF V600 mutation in melanoma. J Transl Med 10, 85.
2.Davies, H., Bignell, G. R., Cox, C., Stephens, P., Edkins, S., Clegg, S., Teague, J., Woffendin, H., Garnett, M. J., Bottomley, W., et al. (2002). Mutations of the BRAF gene in human cancer. Nature 417, 949-954.
3. Smalley, K. S. (2010). Understanding melanoma signaling networks as the basis for molecular targeted therapy. J Invest Dermatol 130, 28-37.
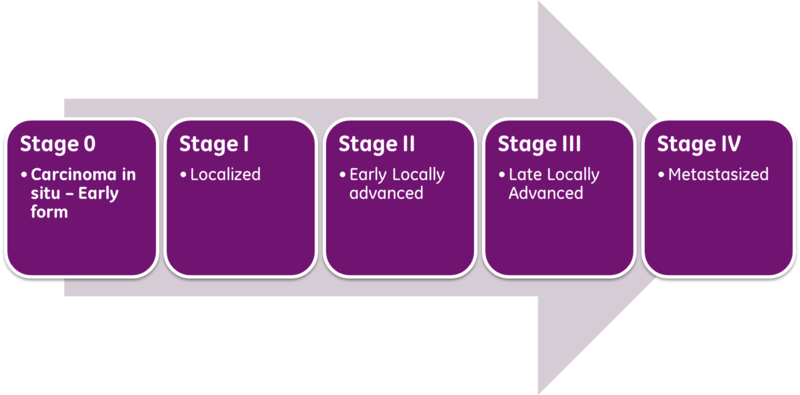
 Classical prognosis of cancer patients utilizes the AJCC/UICC (American Joint Committee on Cancer / International Union Against Cancer) “TNM” classification system, in which T (Tumor) is indicative of primary tumor size and invasion properties, N (Nodes) indicates the extent of tumor invasion into draining and regional lymph nodes, and M (Metastasis), describes the presence and extent of metastatic lesions at diagnosis. The combinations of these parameters are then used to assess a patient’s stage at diagnosis and predict patient outcome. The exact parameter definitions vary for each cancer type.
Classical prognosis of cancer patients utilizes the AJCC/UICC (American Joint Committee on Cancer / International Union Against Cancer) “TNM” classification system, in which T (Tumor) is indicative of primary tumor size and invasion properties, N (Nodes) indicates the extent of tumor invasion into draining and regional lymph nodes, and M (Metastasis), describes the presence and extent of metastatic lesions at diagnosis. The combinations of these parameters are then used to assess a patient’s stage at diagnosis and predict patient outcome. The exact parameter definitions vary for each cancer type.

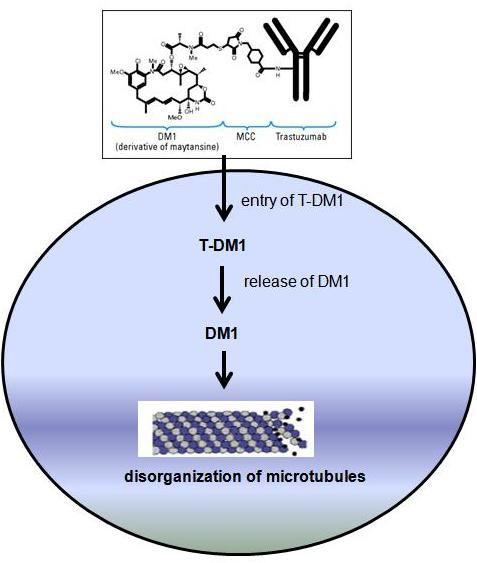
 HER-2 is a 185-kDa orphan transmembrane receptor tyrosine kinase. Dimerization of HER-2 with ligand- bound HER-3 or HER-4 receptor activates signaling pathways inside the cell. Activated HER-2 signaling stimulates cell proliferation and survival via activation of the MAPK and PI3K/Akt/mTOR pathways. Collectively these signaling pathways result in uncontrolled growth of the tumor. Several studies suggested that the overexpression/amplification of HER-2 may lead to the development and progression of pre-malignant breast disease and also tumor metastasis. Therefore, the association of HER-2 in breast cancer as well as its involvement in tumor aggressiveness makes this receptor an appropriate target for tumor-specific therapies. Several strategies have been developed to inhibit HER-2 signaling. These include a tyrosine kinase inhibitor called lapatinib and a recombinant humanized monoclonal antibody called trastuzumab (Herceptin®). In this post I will focus only on trastuzumab mediated therapy in breast cancer. Trastuzumab binds to the extracellular domain of the HER-2 receptor. This inhibits HER-2 signaling via MAPK and PI3K/Akt cascades. In addition, trastuzumab binding also increases membrane localization of the tumor suppressor gene phosphatase and tensin homolog (PTEN), and inhibitor of the PI3K/Aktpathway.
HER-2 is a 185-kDa orphan transmembrane receptor tyrosine kinase. Dimerization of HER-2 with ligand- bound HER-3 or HER-4 receptor activates signaling pathways inside the cell. Activated HER-2 signaling stimulates cell proliferation and survival via activation of the MAPK and PI3K/Akt/mTOR pathways. Collectively these signaling pathways result in uncontrolled growth of the tumor. Several studies suggested that the overexpression/amplification of HER-2 may lead to the development and progression of pre-malignant breast disease and also tumor metastasis. Therefore, the association of HER-2 in breast cancer as well as its involvement in tumor aggressiveness makes this receptor an appropriate target for tumor-specific therapies. Several strategies have been developed to inhibit HER-2 signaling. These include a tyrosine kinase inhibitor called lapatinib and a recombinant humanized monoclonal antibody called trastuzumab (Herceptin®). In this post I will focus only on trastuzumab mediated therapy in breast cancer. Trastuzumab binds to the extracellular domain of the HER-2 receptor. This inhibits HER-2 signaling via MAPK and PI3K/Akt cascades. In addition, trastuzumab binding also increases membrane localization of the tumor suppressor gene phosphatase and tensin homolog (PTEN), and inhibitor of the PI3K/Aktpathway. trastuzumab with taxanes docetaxel (Taxotere®) and paclitaxel (Taxol®) exhibited promising response in HER-2–overexpressing metastatic breast cancer.
trastuzumab with taxanes docetaxel (Taxotere®) and paclitaxel (Taxol®) exhibited promising response in HER-2–overexpressing metastatic breast cancer.
 Mature blood cells are relatively short-lived, and require replenishment from multipotent HSCs. Thus, HSCs must self-renew to generate an adequate pool of HSCs, as well as differentiate to give rise to more mature blood cells. A balance between self-renewal and differentiation ensures that the hematopoietic system can be functionally sustained throughout the lifetime of an adult body. However, as HSCs age, they accumulate DNA damage, often compromising their functionality. DNA damage can be further propagated both to daughter stem cells and downstream lineages, and may increase the risk of developing blood disorders
Mature blood cells are relatively short-lived, and require replenishment from multipotent HSCs. Thus, HSCs must self-renew to generate an adequate pool of HSCs, as well as differentiate to give rise to more mature blood cells. A balance between self-renewal and differentiation ensures that the hematopoietic system can be functionally sustained throughout the lifetime of an adult body. However, as HSCs age, they accumulate DNA damage, often compromising their functionality. DNA damage can be further propagated both to daughter stem cells and downstream lineages, and may increase the risk of developing blood disorders 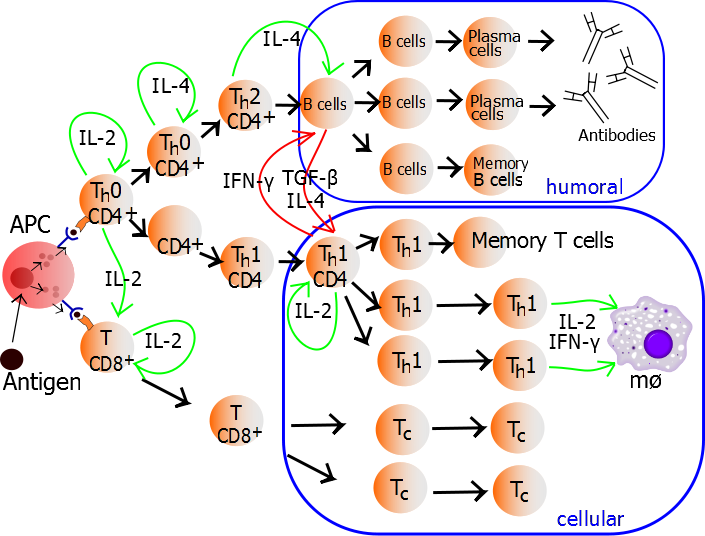
 TH1 cells can be generated and assayed for functions including IFNγ expression in as few as three days. If long term or clonal T cells assays are of interest, cells can be expanded in the presence of IL-2 for 2-3 weeks following single cell cloning.
TH1 cells can be generated and assayed for functions including IFNγ expression in as few as three days. If long term or clonal T cells assays are of interest, cells can be expanded in the presence of IL-2 for 2-3 weeks following single cell cloning. 


 Interferons, including type I (IFNα/β) and type II (IFNγ) are known to be critical for mediating multiple aspects of tumor immunity, by targeting both immune cells for activation, and cancer cells for expression of MHC and genes associated with growth arrest and apoptosis. Thus, expression of cytokines such as IFNγ by immune cells has been shown to be critical in anti-tumor immune responses. IFNγ is one of the major effector cytokines of CD4+ TH1 cells and cytotoxic CD8+ T cells. However, the full effects of IFNγ as well as other TH1 cytokines in mediating anti-tumor effects have not been fully elucidated.
Interferons, including type I (IFNα/β) and type II (IFNγ) are known to be critical for mediating multiple aspects of tumor immunity, by targeting both immune cells for activation, and cancer cells for expression of MHC and genes associated with growth arrest and apoptosis. Thus, expression of cytokines such as IFNγ by immune cells has been shown to be critical in anti-tumor immune responses. IFNγ is one of the major effector cytokines of CD4+ TH1 cells and cytotoxic CD8+ T cells. However, the full effects of IFNγ as well as other TH1 cytokines in mediating anti-tumor effects have not been fully elucidated.