 Natural Killer (NK) cells are a cytotoxic innate immune lymphocyte cell type. In humans, NK cells comprise up to 15% of peripheral blood mononuclear cells (PBMC), and 5-20% of the PBMC lymphocyte population. Several subtypes of NK cells exist in humans. In this post, I will discuss phenotypic properties and markers of NK subtypes present in human PBMC.
Natural Killer (NK) cells are a cytotoxic innate immune lymphocyte cell type. In humans, NK cells comprise up to 15% of peripheral blood mononuclear cells (PBMC), and 5-20% of the PBMC lymphocyte population. Several subtypes of NK cells exist in humans. In this post, I will discuss phenotypic properties and markers of NK subtypes present in human PBMC.
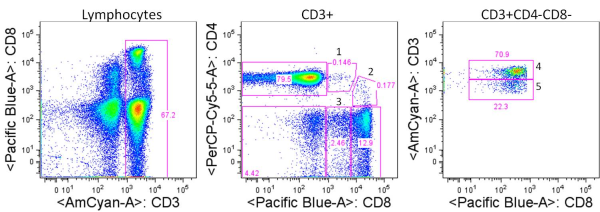
Three subtypes of NK cells are recognized: CD56dim CD16+, CD56brightCD16+/- and CD56– CD16+ NK cells. The CD56dim CD16+ and CD56brightCD16+/- subsets are best studied and are phenotypically classified as a more cytotoxic and a more cytokine producing subset of NK cells, respectively. NK cell activation is mediated by the balance between engagement of activating receptors including NKp46, NKp30, NKp44, NKG2D, CD16, 2B4, NKp80, and DNAM-1, and HLA-I binding inhibitory receptors including killer immunoglobulin-like receptors (KIRs), LIR1/ILT2 and NKG2A/CD94. NK cells can also be activated in response to cytokines such as IL-2, IL-12, IL-15, and IL-18.
CD56dim CD16+ NK cells: This subtype comprises the majority, up to 90%, of PBMC NK cells and is considered the most cytotoxic subset. CD16 is the FCγ receptor III, and can thus bind the FC portion of IgG antibodies and mediate antibody dependant cell-mediated cytotoxicity (ADCC) of antibody-bound target cells. Expression of inhibitory receptors differs among NK subsets, and this subset exhibits lower expression of KIRs and ILT2 but higher expression of NKG2A/CD94 compared with CD56bright NK cells. Expression of granzyme B and perforin is also high in this subset compared with CD56bright NK cells. A recent report by De Maria et. al, demonstrated that this subset does in fact robustly produce cytokines including IFNγ early after activation.
CD56brightCD16+/- NK cells: This subtype comprises up to 10% of NK cells in PBMC, but is the major NK subtype in tissues and secondary lymphoid organs. This subset is conventionally known as the cytokine producing subset of NK cells, and rapidly produces cytokines and chemokines including IFNγ, TNFα, GM-CSF, and RANTES after activation.
Interestingly, in HIV-viremic individuals, a third CD56– CD16+ NK population is significantly expanded in PBMC comprising between 20-55% of NK cells. This population in healthy individuals and aviremic HIV-infected individuals is rare, under 10% of total NK cells. Compared with CD56+ NK cells, the CD56– CD16+ NK cells from HIV-viremic patients exhibited lower expression of activating receptors NKp46, NKp30, and NKp44, lower cytotoxic activity, higher expression levels of inhibitory receptors, and lower expression levels of cytokines including IFNγ, TNFα, and GM-CSF. This subset is also expanded in individuals with chronic HCV infection. Thus, the expansion of this poorly functional NK subset is likely clinically relevant in chronic viral disease.
In summary, these NK populations can be differentiated by expression of CD16 and CD56. Of note, NKT (natural killer-like T) cells can also express these markers along with CD3. Thus, to differentiate these cells from NKT cells, the inclusion of CD3 as a cell identification marker is critical in analysis of these cells by flow cytometry or other methods.
Further Reading:
CD56 negative NK cells: origin, function, and role in chronic viral disease. Björkström NK, Ljunggren HG, Sandberg JK. Trends Immunol. 2010 Nov;31(11):401-6.
The biology of human natural killer-cell subsets. Cooper MA, Fehniger TA, Caligiuri MA. (2001) Trends Immunol 22: 633–640.
Natural killer cell distribution and trafficking in human tissues. Carrega P, Ferlazzo G. Front Immunol. 2012;3:347.
Revisiting human natural killer cell subset function revealed cytolytic CD56(dim)CD16+ NK cells as rapid producers of abundant IFN-gamma on activation. De Maria A, Bozzano F, Cantoni C, Moretta L. Proc Natl Acad Sci U S A. 2011 Jan 11;108(2):728-32.
Natural killer cells in HIV-1 infection: dichotomous effects of viremia on inhibitory and activating receptors and their functional correlates. Mavilio D, Benjamin J, Daucher M, Lombardo G, Kottilil S, Planta MA, Marcenaro E, Bottino C, Moretta L, Moretta A, Fauci AS. Proc Natl Acad Sci U S A. 2003 Dec 9;100(25):15011-6.
Characterization of CD56−/CD16+ natural killer (NK) cells: a highly dysfunctional NK subset expanded in HIV-infected viremic individuals. Mavilio D, Lombardo G, Benjamin J, Kim D, Follman D, et al.. (2005) Proc Natl Acad Sci U S A. 102: 2886–2891.
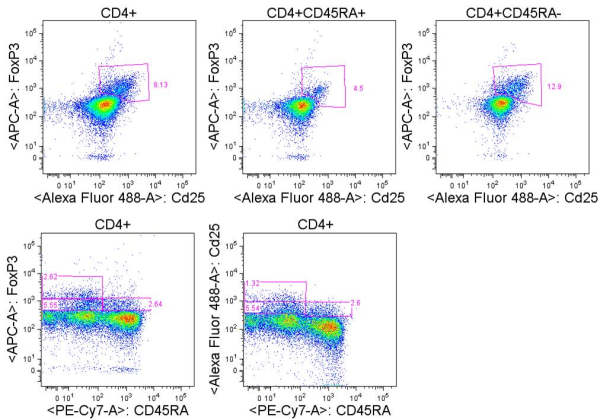
 TH1 cells can be generated and assayed for functions including IFNγ expression in as few as three days. If long term or clonal T cells assays are of interest, cells can be expanded in the presence of IL-2 for 2-3 weeks following single cell cloning.
TH1 cells can be generated and assayed for functions including IFNγ expression in as few as three days. If long term or clonal T cells assays are of interest, cells can be expanded in the presence of IL-2 for 2-3 weeks following single cell cloning. 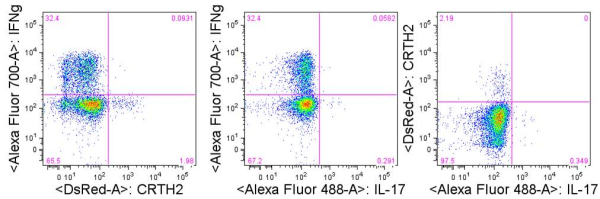

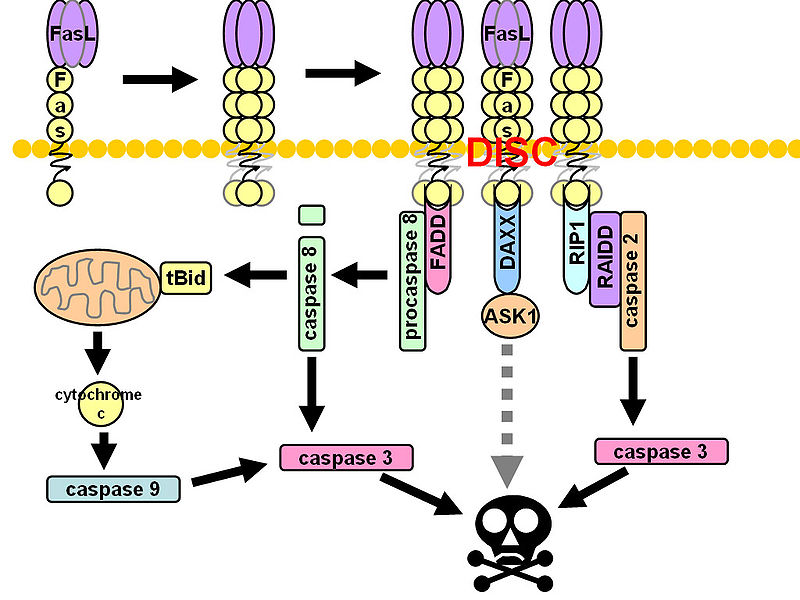
 CD95 (Fas, APO-1, TNFRSF6) is a member of the TNF-receptor superfamily and is best known for its role in mediating activation-induced cell death in activated T cells following binding to its ligand, CD95L/FasL induced on antigen-presenting cells (APCs). However, CD95 can also play additional, non-apoptotic roles in the modulation of T cell function. CD95 ligation has been shown to inhibit TCR signaling and activation of naïve T cells. However, this negative co-stimulatory effect appears to be dose-dependent, as low doses of CD95 agonists had the opposite effect and strongly promoted activation and proliferation of T cells. Like CD71, CD95 expression can be detected by 24 hours following T cell activation and continues to increase over the course of several days.
CD95 (Fas, APO-1, TNFRSF6) is a member of the TNF-receptor superfamily and is best known for its role in mediating activation-induced cell death in activated T cells following binding to its ligand, CD95L/FasL induced on antigen-presenting cells (APCs). However, CD95 can also play additional, non-apoptotic roles in the modulation of T cell function. CD95 ligation has been shown to inhibit TCR signaling and activation of naïve T cells. However, this negative co-stimulatory effect appears to be dose-dependent, as low doses of CD95 agonists had the opposite effect and strongly promoted activation and proliferation of T cells. Like CD71, CD95 expression can be detected by 24 hours following T cell activation and continues to increase over the course of several days.
 There are many strategies for assessing the function of T cells from
There are many strategies for assessing the function of T cells from 


 96-well Plates: Different types of 96-well plates are available for different assay types. There are various surface coatings including tissue-culture treated polystyrene for cell cultures, uncoated, and others. Plates can have various plate bottom geometries and optical characteristics. For instance there are black plates available for light-sensitive assays. For protocols involving volumes larger then 250ul, there are deep-well plates that carry a 2ml volume per well.
96-well Plates: Different types of 96-well plates are available for different assay types. There are various surface coatings including tissue-culture treated polystyrene for cell cultures, uncoated, and others. Plates can have various plate bottom geometries and optical characteristics. For instance there are black plates available for light-sensitive assays. For protocols involving volumes larger then 250ul, there are deep-well plates that carry a 2ml volume per well. Multichannel Vacuums: Companies such as V&P Scientific offer a multitude of multichannel vacuum manifolds that fit plates of different depths for removing supernatant from wells via vacuum apparatus. Often these will be the proper length such that they don’t touch the well bottom and work well with removing buffers from centrifuged PBMC cell cultures, such as during washing steps for flow-cytometry.
Multichannel Vacuums: Companies such as V&P Scientific offer a multitude of multichannel vacuum manifolds that fit plates of different depths for removing supernatant from wells via vacuum apparatus. Often these will be the proper length such that they don’t touch the well bottom and work well with removing buffers from centrifuged PBMC cell cultures, such as during washing steps for flow-cytometry.